HIALOPLASMA, CITOESQUELETO
Y ESTRUCTURAS NO MEMBRANOSAS I
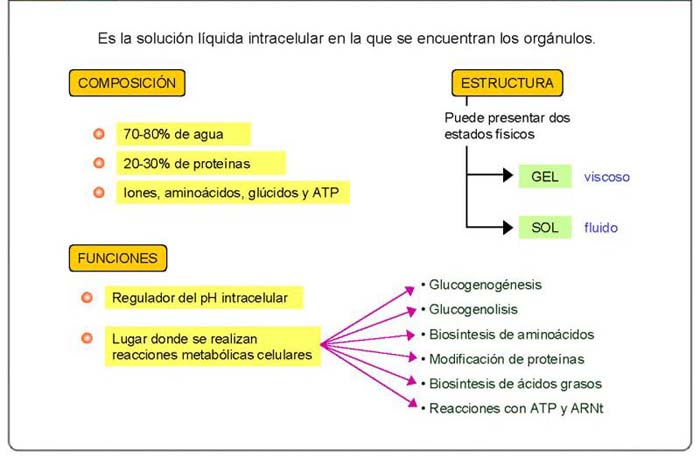
HIALOPLASMA O CITOSOL
Es el medio acuoso del citoplasma en el que se encuentran inmersos los orgánulos celulares.
Citosol + orgánulos = Citoplasma.
Composición70 - 80% agua, 30 - 20% proteínas (enzimas), mol. pequeñas: glúcidos, aá, iones, etc.
Debido a esta gran concentración de proteínas, el citosol es un gel viscoso organizado por las fibras citoesqueléticas.
La mayoría de las proteínas en el hialoplasma son enzimas que catalizan un gran número de reacciones del metabolismo celular.
Además, en el citosol de muchas células se almacenan sustancias de reserva en forma de gránulos, denominados inclusiones, que no están rodeados por una membrana. Así, las células musculares y los hepatocitos contienen gránulos citosólicos de glucógeno y los adipocitos contienen grandes gotas de grasa, que pueden llegar a ocupar casi todo el citosol.
EstructuraGel <> Sol: motilidad celular.
Movimientos citoplasmáticos y paso de sol a gel y viceversa del hialoplasma en una célula para desplazarse mediante pseudópodos
Funciones
Regulador de pH.
- Glucolisis (degradación de la glucosa) y gluconeogénesis (biosíntesis de azúcares)
- Glucogenolisis y Glucogenogénesis
- Biosíntesis y degradación de aminoácidos
- Biosíntesis de nucleótidos
- Biosíntesis de ácidos grasos
- Reacción de aminoácidos y ARNt (ATP)
CITOESQUELETO
El interior de la célula eucariota no es una masa amorfa y gelatinosa donde están diseminados al azar el núcleo y el resto de los orgánulos. Por el contrario, posee una organización interna establecida por una serie de filamentos proteicos que forman un entramado dinámico y se extienden a través del citoplasma, sobre todo, entre el núcleo y la cara interna de la membrana celular, aunque también los hay intranucleares. A esta matriz proteica y fibrosa se la denomina citoesqueleto.
Armazón tridimensional de filamentos proteicos, entramado reticular +/- denso que se extiende por todo el hialoplasma de las células eucariotas.
Es una estructura dinámica: se ensambla y se reorganiza continuamente según las necesidades celulares
Citoesqueleto
Funciones
Sus funciones son particularmente importante en las células animales, donde no existe una pared celular que de consistencia a las células.
- Proporcionar soporte interno a las estructuras y orgánulos celulares
- Mantener la morfología celular
- Organización interna
- Facilitar la motilidad célular (formación de pseudópodos, cilios y flagelos) y de transporte intracelular
- Intervenir en la división celular (huso acromático)
Componentes
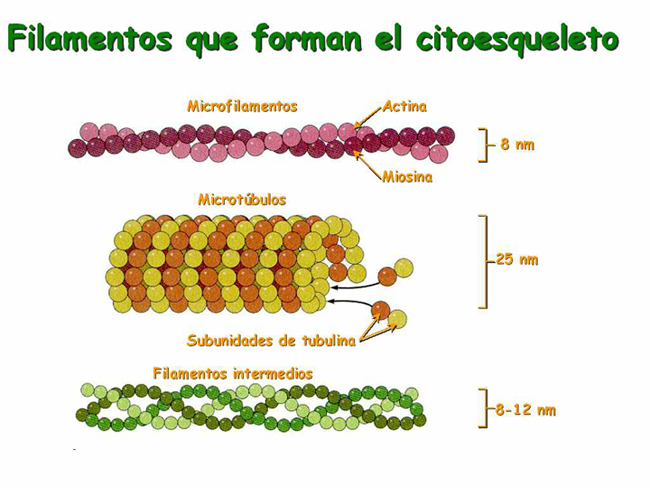
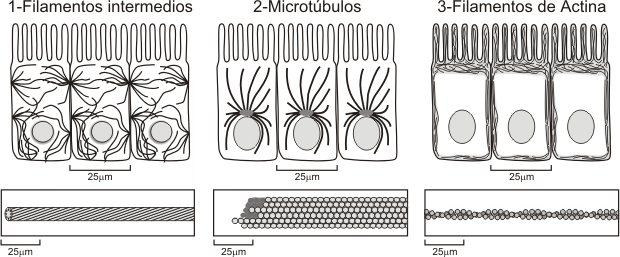
Tres tipos de componentes:- Filamentos de actina o microfilamentos (7-8 nm)
- Filamentos intermedios (10 nm)
- Microtúbulos (25 nm)
Además, estos sistemas de filamentos (microfilamentos, filamentos intermedios y microtúbulos), se mantienen unidos a la membrana plasmática y a los orgánulos celulares mediante proteínas accesorias.
Las proteínas accesorias cumplen distintas funciones y se las clasifica en:
- Proteínas reguladoras: regulan los procesos de alargamiento (polimerización) y acortamiento (despolimerización) de los filamentos principales.
- Proteínas ligadoras: conectan los filamentos entre si y con distintas estructuras celulares
- Proteínas motoras: sirven para la motilidad, contracción y cambios de forma celulares. También trasladan macromoléculas y organoides de un punto a otro del citoplasma.
Microfilamentos (o filamentos de actina)
Estructuras flexibles en forma de haces, redes.
En las células animales se encuentran normalmente localizados cerca de la membrana plasmática.
Es el componente del citoesqueleto más dinámico (el que más se modifica).
Distribución de los filamentos de actina en la célula:
1. Filamentos Transcelulares (atraviesan el citoplasma en todas las direcciones).
2. Filamentos Corticales (por debajo de la membrana plasmática)
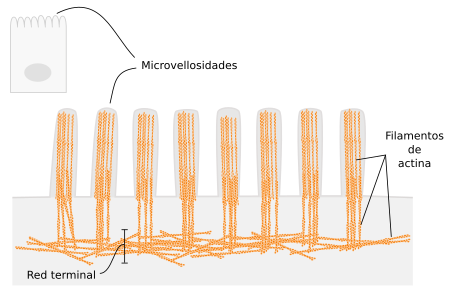
Los filamentos de actina poseen entre 3 y 7 nm de diámetro. Es el valor más pequeño dentro de los filamentos que componen el citoesqueleto, por ello también se denominan microfilamentos.
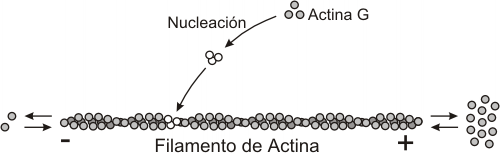
Formados por una proteína globular, actina, que en general constituye entre el 5 y el 10% del total de las proteínas celulares. Puede presentarse de dos formas:- Actina no polimerizada (actina G). Se trata de una proteína globular asociada a la proteína profilina que evita su polimerización. Se encuentra disuelta en el citosol y representa la mitad de la actina de la célula y polimeriza microfilamentos cuando son necesarios.
- Actina polimerizada o filamentosa (actina F). En presencia de ATP, la actina G se polimeriza formando largas hélices dobles, denominadas actina F, o actina filamentosa. Consta de dos filamentos enrollados entre sí (doble hélice dextrógira) de actina globular (actina G).
Una de las propiedades de la actina F es su polaridad +/-, es decir, presentan un comportamiento diferente entre sus dos extremos:
- el extremo más se denomina así porque en él predomina la polimerización, adición de nuevas moléculas de actina, respecto a la despolimerización, mientras que,
- en el extremo menos predomina la despolimerización.
Polimerización y despolimerización de los filamentos de actina.
(a) actina G, (b) nucleación, (c) polimerización y despolimerización
El mecanismo de crecimiento y acortamiento de la longitud de los filamentos de actina es por polimerización y despolimerización, respectivamente, de monómeros de actina.
En las células se crean y se destruyen filamentos de actina continuamente.
La actina F puede aparecer asociada a otras proteínas que modifican sus propiedades:
- Proteínas estructurales: permiten la unión o anclaje de los filamentos de actina, principalmente a proteínas de la membrana plasmática.
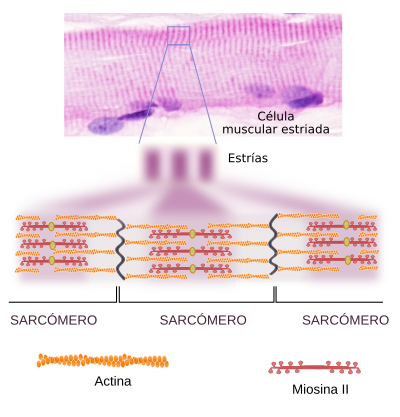
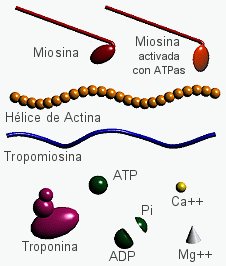
Los filamentos de actina abundan debajo de la mb plasmática donde forman una estructura reticular densa denominada córtex celular, responsable del mantenimiento de la morfología celular y de la ordenación y desplazamiento de las proteínas en el plano de la membrana plasmática, a través de las interacciones con las proteínas estructurales.- Proteínas reguladoras: La más importante es la miosina, que participa en la contracción muscular al permitir que la actina se desplace sobre ella.
Funciones de los filamentos de actina
Estructuras dinámicas: Relacionadas con la motilidad
- Contracción muscular (haces musculares): Las proteínas motoras que se asocian con la actina para producir el movimiento muscular son del tipo de las miosinas.
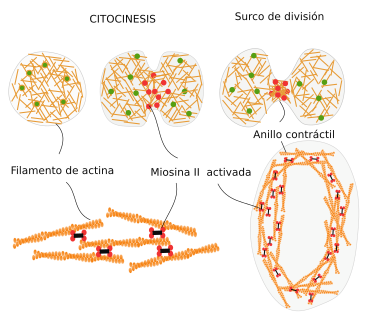
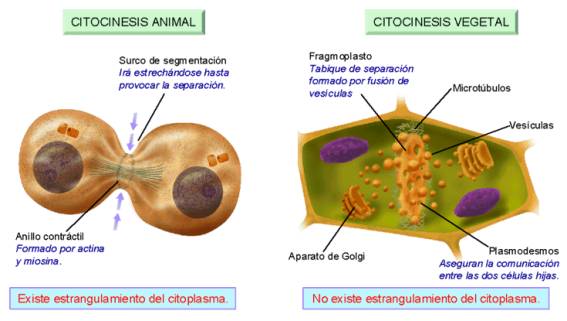
- Formación del anillo contráctil durante la división del citoplasma (citocinesis) en la división celular de las células animales. El estrangulamiento final del citoplasma durante el proceso de división celular se produce gracias a un anillo de actina, que, ayudado por la miosinas, va estrechando su diámetro progresivamente hasta la separación completa de los dos citoplasmas de las células hijas.
Citocinesis en células animales (con participación de microfilamentos)
y en células vegetales (no se forma anillo contráctil)
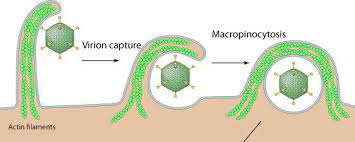
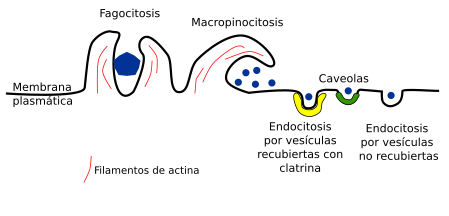
- Participación en los procesos de endocitosis, fagocitosis: La formación y escisión de vesículas en la membrana plasmática no se realiza si se impide la polimerización de los filamentos de actina. La emisión de las expansiones celulares que engloban a las moléculas que van a ser fagocitadas dependen de la polimerización de de filamentos de actina.
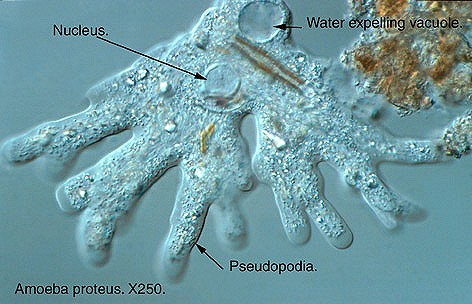
- Formación de pseudópodos y generación de corrientes citoplasmáticas: Las células no nadan, se desplazan arrastrándose por el medio que las rodea y ello se hace por un mecanismo de reptación, como ocurre en las células embrionarias durante el desarrollo, en el desplazamiento de las amebas, en la invasión de los linfocitos de los tejidos infectados
Ameba y pseudópodos
Estructuras rígidas:
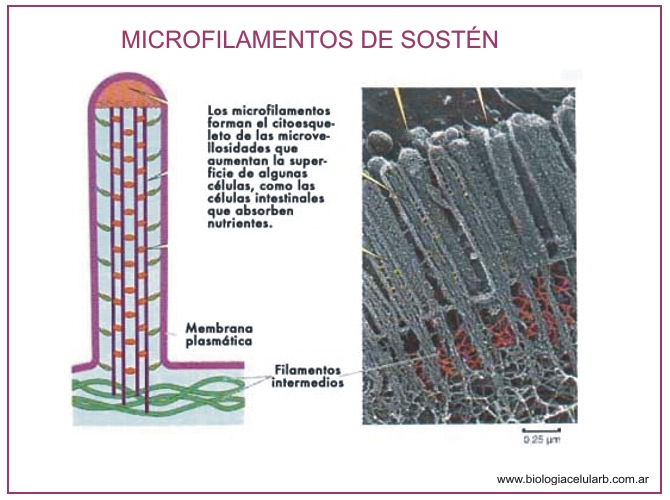
- Formación de las microvellosidades intestinales: Las microvellosidades son estructuras estables que permiten a la célula aumentar enormemente la superficie de su membrana plasmática y aparecen en las células epiteliales como las del tubo digestivo, donde se aumenta enormemente la superficie de absorción.
Son tubos largos y relativamente rígidos, de unos 25 nm de diámetro, con una luz central.
El sistema de microtúbulos de las células animales se forma principalmente a partir del centrosoma y desde este se extienden a todas las partes de la célula.
Distribución de los microtúbulos en la célula
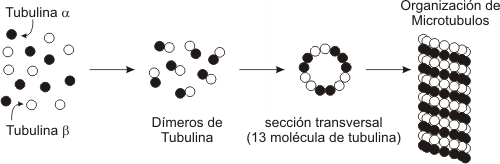
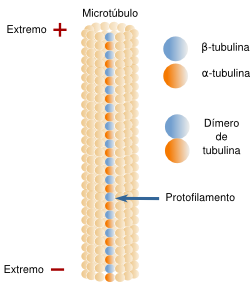
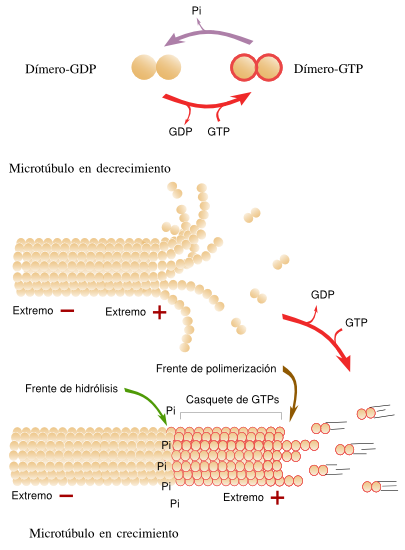
Las paredes de los microtúbulos están formados por unas subunidades proteicas globulares denominadas tubulinas. Éstas se asocian en dímeros compuestos por dos tipos de tubulinas: α y β.
Ambas proteínas se alinean ordenadamente, en paralelo, formando heterodímeros, en filas longitudinales que se denominan protofilamentos.
Un microtúbulo tipo contiene trece protofilamentos que forman un cilindro.
Polimerización de la tubulina a partir de las tubulinas alfa y beta
Esquema de la organización de los dímeros de tubulina en un protofilamento que forma parte de un microtúbulo. Nótese que la α-tubulina está orientada hacia el extremo menos y la β-tubulina hacia el extremo más.
Los microtúbulos al igual que los filamentos de actina, presentan polaridad: se alargan o polimerizan agregando dímeros de tubulina en uno de sus extremos (extremo positivo), mientras que por el otro se acortan o despolimerizan eliminando dímeros (extremo negativo).
Extremos + y - de un microtúbulo
Los microtúbulos son estructuras también muy dinámicas que pueden formarse o destruirse según las necesidades de las células. Crecen y se acortan constantemente.
Son más flexibles, pero más duros que los microfilamentos de actina.
Funciones de los microtúbulos
Microtúbulos estables: Función estructural
- Formación de cilios y flagelos.
- Organización celular en torno al centrosoma
- Mantenimiento de la forma: eritrocitos.
Microtúbulos dinámicos o cambiantes: Función dinámica
- Transporte intracelular. Distribución de orgánulos, vesículas de secreción y transporte intracelular de sustancias. Capacidad de movimientos.
- Formación del huso mitótico.
- Movimiento celular.
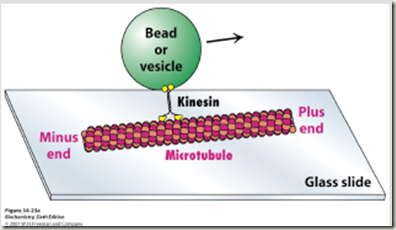
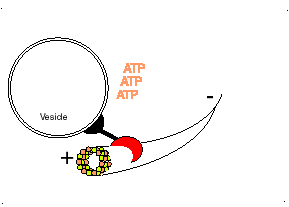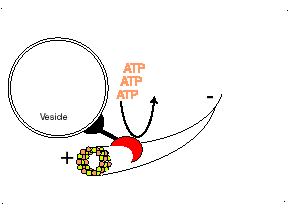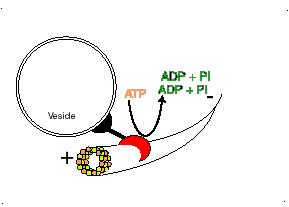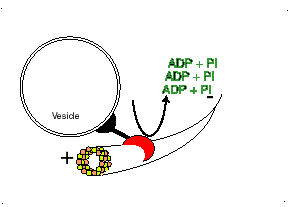
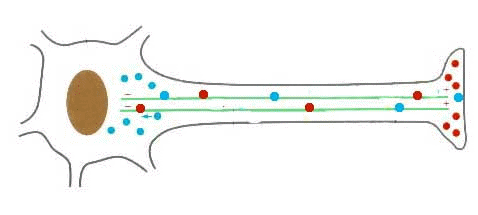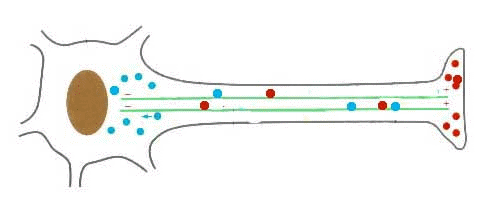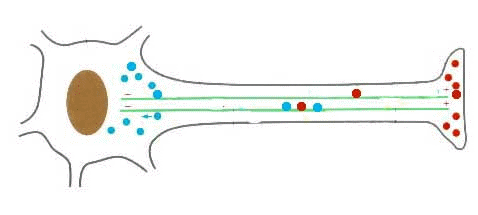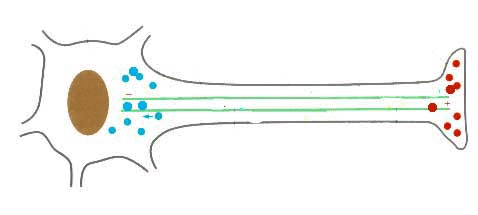
Los microtúbulos participan en numerosos procesos como la organización intracelular o la división celular, gracias a la colaboración de proteínas motoras: dineínas y quinesinas, las cuales se desplazan por el microtúbulo en direcciones opuestas: las quinesinas hacia el extremo más y las dineínas hacia el extremo menos.
El movimiento conlleva gasto de ATP.
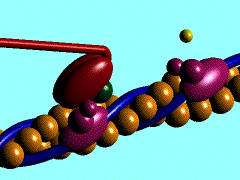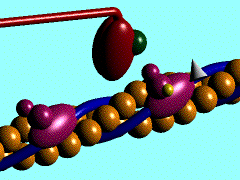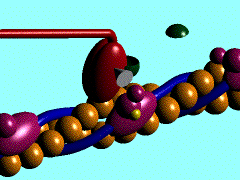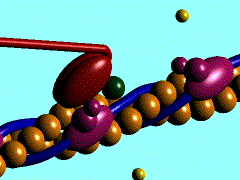
Proteína kinesina transportando una vesícula utilizando un microtúbulo
Proteína dineína transportando una vesícula utilizando un microtúbulo
Las DINEINAS se mueven hacia el extremo NEGATIVO del microtúbulo (o sea hacia el centrosoma), las KINESINAS se mueven hacia el extremo POSITIVO.
ver movimiento de la dineina por los microtúbulos
ver vídeo del tráfico de vesículas por la célula
ver animación de las proteínas motoras: kinesina y dineina
Formados por diferentes proteínas fibrosas, son exclusivos de las células animales.
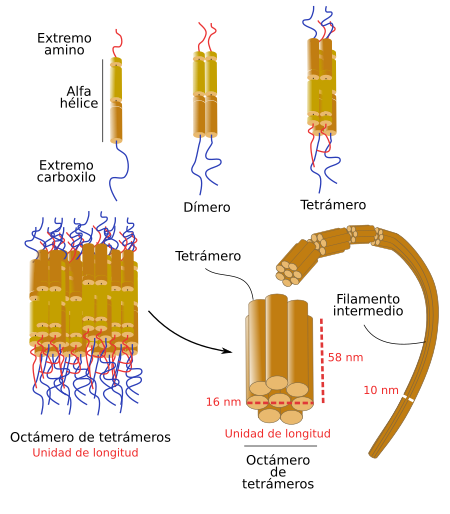
Tienen unos 10-12 nm de diámetro.
Presentan una gran resistencia a las tensiones mecánicas y su principal misión es permitir a las células soportar tensiones mecánicas cuando son estiradas. Son flexibles y resistentes, dos propiedades óptimas para soportar las tensiones mecánicas. Se ha estimado que pueden estirarse entre un 250 y un 350 % de su longitud inicial cuando se someten a fuerzas de tensión.
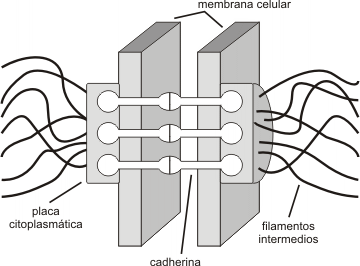
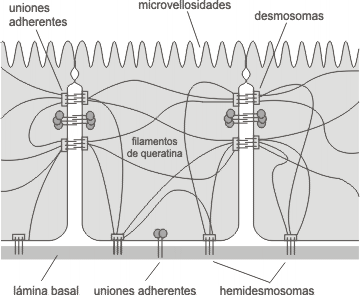
Son los responsables de mantener la integridad celular puesto que funcionan a modo de cables intracelulares que se enganchan a complejos de unión como los desmosomas y los hemidesmosomas, lo que permite la cohesión entre células contiguas y por tanto la cohesión celular.
Estructura de un desmosoma
Representan los componentes más estables del citoesqueleto
Los filamentos intermedios forman redes que conectan la membrana plasmática con la envoltura nuclear, formando una red continua a su alrededor. Una red similar se encuentra en la cara interna de la envoltura (lámina nuclear)
También se han encontrado filamentos intermedios en el núcleo donde forman la lámina nuclear,
un entramado que da forma y aporta cohesión a la envuelta nuclear.
A diferencia de los microfilamentos y microtúbulos, los filamentos intermedios al agruparse pierden polaridad, por lo tanto no presentan extremo + y extremo -
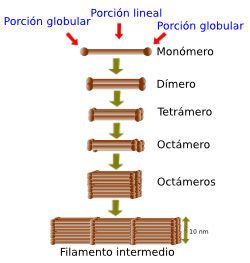
Están formados por monómeros con dos dominios: cabezas o zonas globulares y zonas intermedias.
Esquema del ensamblaje de los filamentos intermedios a partir de monómeros.
Los filamentos intermedios pueden ser específicos en algunas células. Los principales tipos de filamentos intermedios son:
- Neurofilamentos: en los axones de las neuronas
- Filamentos de queratina o tonofilamentos: formados por queratina, aparecen en células epiteliales, principalmente en los desmosomas.
- Filamentos de vimentina: se encuentran en las células del tejido conjuntivo y vasos sanguíneos.
- Filamentos de desmina: presentes en las células musculares
Uniones de anclaje de los filamentos intermedios en células epiteliales
Funciones de los filamentos intermedios
Función estructural
- Resistencia mecánica, se unen a desmosomas.
- Forma celular
Distribución de los diferentes tipos de componentes del citoesqueleto en la célula
ver vídeo-animación citoesqueleto
ver vídeo ¿qué es el citoesqueleto?