ORGÁNULOS ENERGÉTICOS II
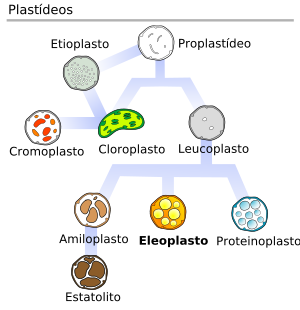
PLASTOS o PLASTIDIOS
Orgánulos característicos y exclusivos de las células eucariotas vegetales.
Forma y tamaño variable. Visibles al microscopio óptico.
Delimitados por una doble membrana. Presentan ADN circular.
Todos los plastos proceden de proplastidios, que son orgánulos pequeños (0,5 - 1 micras de diámetro) e indiferenciados, presentes en células en división de las raíces y los brotes de las plantas. En función de las necesidades de la planta, los proplastidios darán lugar a distintos tipos de plastidios maduros, que pueden transformarse de un tipo a otro.
El conjunto de plastos de una célula se denomina plastidoma.
Tipos de plastos
Existen diferentes tipos de plastos que se diferencian en su estructura y función.
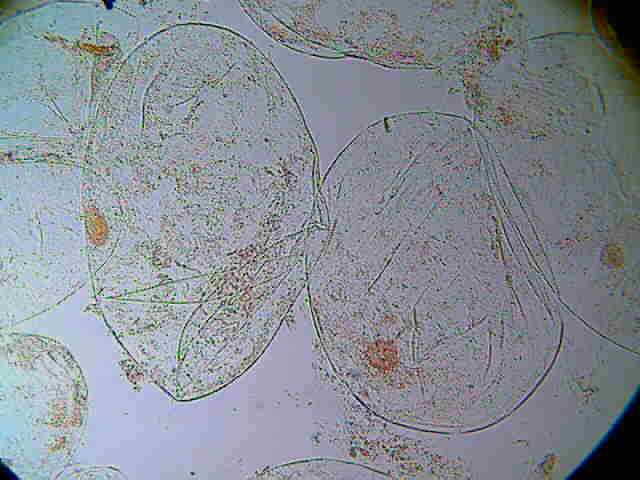
Cloroplastos
Fotosintéticamente activos.
Son los plastos más abundantes y de mayor importancia biológica.
Son de color verde debido a que fabrican y almacenan la clorofila (los vemos más adelante).
Cloroplastos al M. óptico
Los feoplastos (color pardo debido a la fucoxantina) y los rodoplastos (rojos) propios de algunos tipos de algas también son fotosintéticamente activos.
En los rodoplastos de algas rodofíceas, la clorofila está enmascarada por carotenoides de color rojo (ficoeritrina-roja y ficocianina azul) que proporciona el color rojo a dichas algas.
Alga roja Calliblepharis cilliata
Fotosintéticamente inactivos.
No poseen clorofilas. Sintetizan y almacenan pigmentos.
Poseen pigmentos como carotenos (amarillos o anaranjados), xantofilas (amarillentas), licopeno (rojo), etc. a los que deben su color.
Se encuentran en raíces como la zanahoria, frutos como el tomate y pimiento o pétalos de algunas flores como las del género Tropaeolum.
Durante la maduración de los frutos, como el tomate, los cromoplastos se originan a partir de cloroplastos.
A lo largo de este proceso se produce la síntesis de pigmentos carotenoides acompañada de la modificación o desaparición del sistema de tilacoides y de la descomposición de la clorofila.
Cromoplastos de tomate al microscopio óptico
La diferenciación de un cromoplasto no es un fenómeno irreversible. Un ejemplo: en la parte superior de las raíces de la zanahoria expuestas a la luz, los cromoplastos pueden diferenciarse en cloroplastos perdiendo los pigmentos y desarrollando tilacoides, y adquiriendo un color verde.
Fotosintéticamente inactivos.
Estos plastos son incoloros ya que carecen de pigmentos
Se localizan en las células vegetales de órganos no expuestos a la luz, tales como raíces, tubérculos, semillas y órganos que almacenan almidón.
Amacenan distintas sustancias de reserva como almidón (Amiloplastos), aceites (Oleoplastos o elaioplastos), proteínas (Proteoplastos)
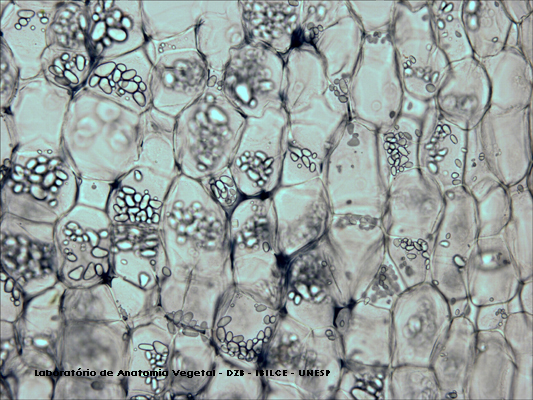
- Amiloplastos: almacenan almidón que se forma en los cloroplastos a través de la fotosíntesis, después es hidrolizado y se resintetiza como almidón de reserva en los amiloplastos. Poseen forma variada: pueden ser esféricos, ovales o alargados, y normalmente muestran una deposición en capas alrededor de un punto. Se tiñen de color azul-negro con compuestos yodados (lugol)
Células vegetales con amiloplastos al microscopio óptico
- Proteinoplastos: acumulan proteínas en forma de cristales o filamentos. Frecuentes en los elementos cribosos del floema.
- Oleoplastos o elaioplastos: acumulan lípidos como los aceites. Son frecuentes en la pulpa de la aceituna y en los cotiledones del girasol y cacahuete.
CLOROPLASTOS
Orgánulos encargados de la realización de la fotosíntesis, en células eucariotas vegetales.
Son los plastos más abundantes y de mayor importancia biológica.
Son de color verde debido a que fabrican y almacenan la clorofila.
En presencia de luz, los cloroplastos se desarrollan a partir de proplastidios.
Si las plantas se mantienen en la oscuridad, el desarrollo de los proplastidios se detiene en un estado intermedio denominado etioplasto, que no contiene clorofila y en el que se desarrolla una estructura semicristalina de membranas tubulares.
Si las plantas que han crecido en la oscuridad, se exponen a la luz, los etioplastos continúan su desarrollo hasta convertirse en cloroplastos.
Los cloroplastos viejos, amarillos por la presencia de carotenoides y que aparecen en las hojas en el otoño, se llaman gerontoplastos y contituyen la última etapa (irreversible) en el desarrollo de los plastos.
Distribución y morfología
Células eucariotas vegetales fotosintéticas: parénquimas clorofílicos.
nº variable (en general, de 1 - 40) dependiendo del tipo de tejido vegetal.
Tamaño: 2 a 6 µm de diámetro, 5 a 10 µm de largo.
Forma muy variable: Normalmente ovoides, copa, hélice, estrellados, etc.
Localización: frecuentemente agrupados cerca del núcleo o hacia la pared celular, aunque también homogéneamente por todo el citoplasma.
Poseen capacidad de movimiento gracias a las corrientes citoplasmáticas o ciclosis.
Se multiplican, al igual que las mitocondrias, por división (bipartición): alargamiento del plasto y constricción en su parte central.
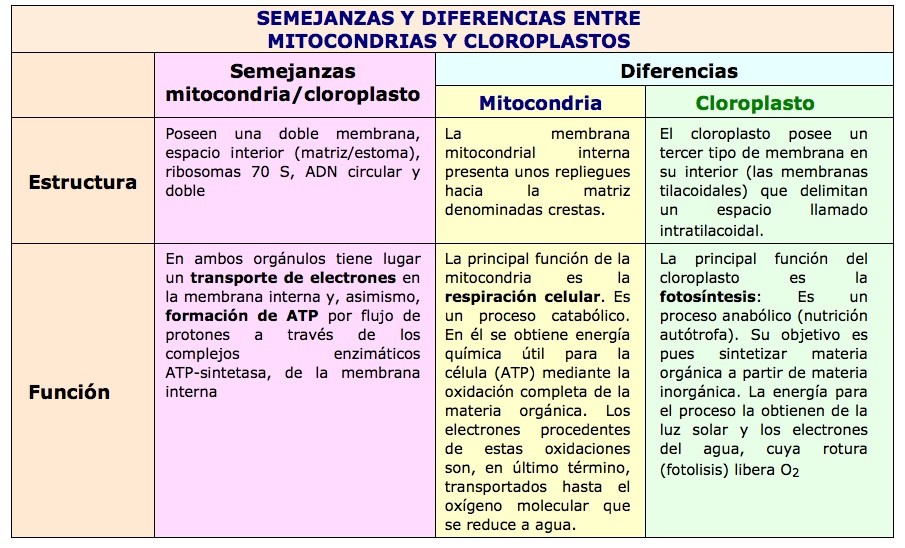
Similares a las mitocondrias en la función (en tamaño son mucho más grandes): ambos orgánulos generan energía, contienen su propio genoma y se replican por división.
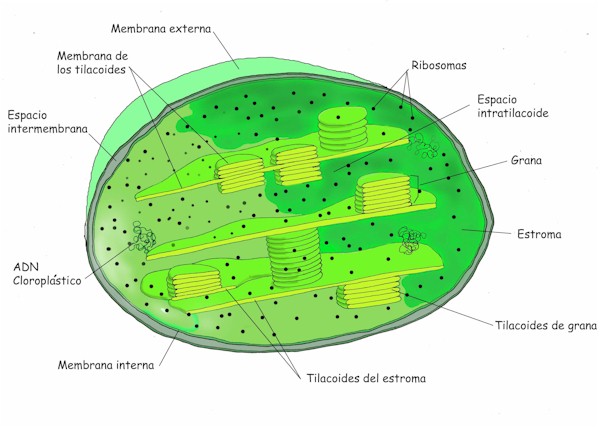
Estructura-ultraestructura del cloroplasto
Delimitados por una doble membrana:
Membrana (cloroplástica) externa: muy permeable, similar a la plasmática y a la membrana externa de las mitocondrias. Porinas.
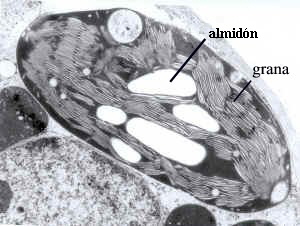
- Tilacoides de grana, sáculos membranosos aplanados que tienden a formar apilamientos a modo de monedas denominados grana. En el estroma puede haber unos 40-60 grana y cada uno de ellos una docena de tilacoides
Las membranas de los tilacoides contienen los pigmentos fotosintéticos, principalmente clorofilas y carotenoides, la cadena fotosintética de transporte de electrones y la ATP sintetasa.
En la cara estromal:
- Fotosistemas, cadena de transporte electrónico
- Partículas elementales F1: Complejo ATP-sintetasas.
Esquema resumido de las moléculas que participan en la fase luminosa de la fotosíntesis. Todas están asociadas a la membrana de los tilacoides. Los protones se bombean al interior del tilacoide, mientras que el ATP y NADPH quedan en el estroma del cloroplasto.
Estructura de la ATP-sintetasa del cloroplasto
- Tilacoides del estroma: conectan a los tilacoides de diferentes grana entre ellos formando una red de cavidades. Paralelos al eje mayor. 12% de pigmentos.
- Espacio tilacoidal o intratilacoidal (intralamelar o lumen): situado en el interior de los tilacoides que mantiene unas condiciones de pH ácido.
Microfotografía electrónica de un corte de parte de un cloroplasto,
donde se observan las granas, apilamientos con un número variable de sacos interconectados, formando una red membranosa interna: espacio tilacoidal.
- Enzimas implicados en la fotosíntesis: Enzimas de la fase oscura de la fotosíntesis: De ellos, la más abundante es la ribulosa bifosfato carboxilasa oxigenasa (Rubisco: fijación del C), que puede llegar a representar la mitad de las proteínas del cloroplasto.
- ADN de doble cadena y circular. El genoma de los cloroplastos es mayor que el mitocondrial. Contiene información genética que codifica distintos tipos de ARN y algunas proteínas de los complejos enzimáticos que participan en la fotosíntesis.
La existencia de ADN cloroplástico confiere a los cloroplastos cierta autonomía respecto al núcleo celular, aunque no los hace totalmente autosuficientes ya que necesitan numerosas proteínas codificadas por el ADN nuclear que posteriormente son importadas a su interior.
El ADN cloroplástico codifica unas 30 proteínas ribosómicas que representan aproximadamente un tercio de las proteínas del cloroplasto.
- Ribosomas (plastorribosomas): encargados de la síntesis de las proteínas propias del cloroplasto. Son más pequeños que los ribosomas ctitoplasmáticos y similares a los bacterianos. Coeficiente de sedimentación 70s
Ultraestructura del cloroplasto al M.E.T y esquema
- Enzimas que regulan y controlan la replicación, transcripción y traducción del material genético del cloroplasto.
- Sustancias diversas: principalmente gránulos de almidón y gotas lipídicas (plastoglóbulos).
ver imágenes de cloroplastos al MET
Funciones de los cloroplastos
- Fase luminosa: Energía luminosa>>> ATP, NADPH
- Fase oscura: Fijación de CO2
- Síntesis de glúcidos.
ver animación viaje al interior de un cloroplasto
ver video cloroplasto y fotosíntesis
ORIGEN DE LAS MITOCONDRIAS Y CLOROPLASTOS
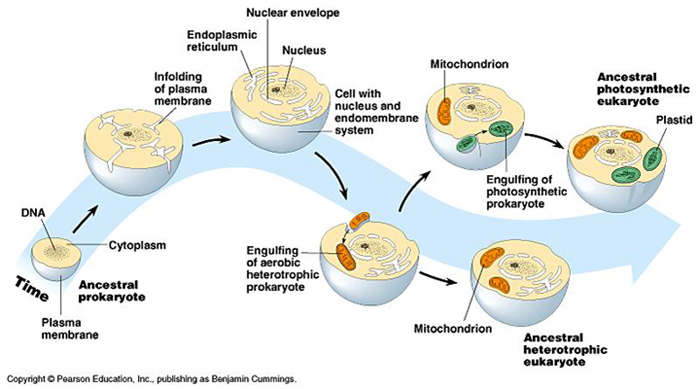
La Teoría endosimbiótica o endosimbiosis serial, formulada a principios del siglo XX y desarrollada por Lynn Margulis, trata de explicar el origen de las mitocondrias y cloroplastos mediante una serie de incorporaciones simbiogenéticas.
Lynn Margulis
Actualmente a esta Teoría se le han ido añadiendo algunos cambios a partir de los nuevos descubrimientos y puede resumirse así:
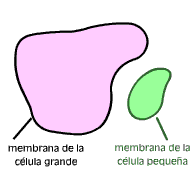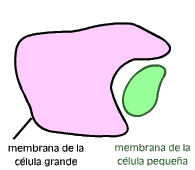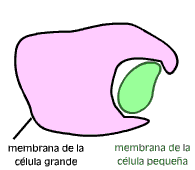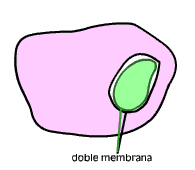
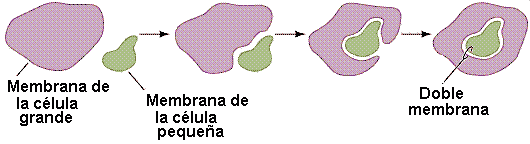
El primer Eobionte fue probablemente una célula procariota anaerobia y heterótrofa. Este organismo desarrollo un sistema de endomembranas que confinó al ADN en un núcleo, separado del resto del citoplasma por una doble membrana a la vez que desarrollo la fagocitosis como una forma de incorporar los materiales que necesitaba.
A través de la fagocitosis, esta primitiva célula eucariota pudo llevar a cabo una serie de incorporaciones simbiogenéticas cuyo resultado fue la aparición de mitocondrias y cloroplastos (en algunas de ellas).
Primera incorporación simbiogenética: origen de las mitocondriasHace unos 1.500 m.a., la primitiva célula procariota fagocitó bacterias aeróbicas que en vez de ser digeridas, se quedaron en el citoplasma, donde se reprodujeron libremente hasta dar lugar a las mitocondrias.
ver vídeo incorporación de las mitocondrias
(Teoría endosimbiótica serial)
Segunda incorporación simbiogenética: origen de los cloroplastos
Mediante un proceso similar, unos 100 m.a. después de la aparición de las mitocondrias, en algunas de esas células, se produjo un segundo suceso. En algunas de esas primitivas células, ya con mitocondrias, se incorporaron mediante fagocitosis, cianobacterias (fotosintéticas oxigénicas) que derivarían a los actuales cloroplastos.
Estas células darían lugar a las células eucariotas vegetales actuales.
Teoría endosimbiótica de la célula eucariota
Tercera incorporación simbiogenética
Los cloroplastos de los Cromistas (un grupo dentro de los Protoctistas) poseen hasta cuatro membranas y se formarían a partir de incorporaciones posteriores endosimbióticas de células eucariotas que ya poseían cloroplastos.
ver vídeo Teoría endosimbiótica serial
Pruebas de la Teoría endosimbiótica
La Teoría endosimbiótica, actualmente aceptada por casi la totalidad de la comunidad científica, se apoya entre otras pruebas en:
- El tamaño de las mitocondrias es similar al tamaño de algunas bacterias.
- Las mitocondria y los cloroplastos contienen ADN bicatenario circular cerrado - al igual que los procariotas- mientras que el núcleo eucariota posee varios cromosomas bicatenarios lineales.
- Están rodeados por una doble membrana, lo que concuerda con la idea de la fagocitosis: la membrana interna sería la membrana plasmática originaria de la bacteria, mientras que la membrana externa correspondería a aquella porción que la habría englobado en una vesícula.
- Las mitocondrias y los cloroplastos se dividen por fisión binaria al igual que los procariotas (los eucariotas lo hacen por mitosis). En algunas algas, tales como Euglena, los plastos pueden ser destruidos por ciertos productos químicos o la ausencia prolongada de luz sin que el resto de la célula se vea afectada. En estos casos, los plastos no se regeneran.
- En mitocondrias y cloroplastos los centros de obtención de energía se sitúan en las membranas, al igual que ocurre en las bacterias. Por otro lado, los tilacoides que encontramos en cloroplastos son similares a unos sistemas elaborados de endomembranas presentes en cianobacterias.
- En general, la síntesis proteica en mitocondrias y cloroplastos es autónoma.
- Algunas proteínas codificadas en el núcleo se transportan al orgánulo, y las mitocondrias y cloroplastos tienen genomas pequeños en comparación con los de las bacterias.Esto es consistente con la idea de una dependencia creciente hacia el anfitrión eucariótico después de la endosimbiosis. La mayoría de los genes en los genomas de los orgánulos se han perdido o se han movido al núcleo. Es por ello que transcurridos tantos años, hospedador y huésped no podrían vivir por separado.
- En mitocondrias y cloroplastos encontramos ribosomas 70s, característicos de procariotas, mientras que en el resto de la célula eucariota los ribosomas son 80s.
- El análisis del ARNr 16s de la subunidad pequeña del ribosoma de mitocondrias y plastos revela escasas diferencias evolutivas con algunos procariotas.
- Una posible endosimbiosis secundaria (es decir, implicando plastos eucariotas) ha sido observado por Okamoto e Inouye (2005). El protista heterótrofo Hatena se comporta como un depredador e ingiere algas verdes, que pierden sus flagelos y citoesqueleto, mientras que el protista, ahora un anfitrión, adquiere nutrición fotosintética, fototaxia y pierde su aparato de alimentación.
ver animación teoría endosimbiotica de la célula eucariota
ver animación teoría endosimbiotica de la célula eucariota II
ver animación teoría endosimbiotica de la célula eucariota III
ver más información sobre la Teoría Endosimbiótica de la célula eucariota (Wikipedia)