PROCESOS CATABÓLICOS V
CATABOLISMO DE LIPIDOS (ACILGLICEROLES o GRASAS)
En los animales superiores las grasas tienen una gran importancia como combustibles orgánicos ya que poseen un alto valor calórico. La degradación de 1 g de grasa puede proporcionar hasta 9 Kcal mientras que como vimos en el catabolismo de los azúcares, cada gramo de glucosa, 4 Kcal.
Los lípidos pueden proceder de:
- los triacilglicéridos ingeridos
- de los almacenados en tejidos de reserva como el adiposo
- de los fabricados por el hígado a partir del excedente de glúcidos en la dieta.
Las grasas constituyen la principal reserva energética del organismo debido a:
- Sus átomos de carbono están casi totalmente reducidos en comparación con los de los azúcares o aminoácidos, por lo que su oxidación proporciona más ATP
- Al ser insolubles en agua no se hidratan y se pueden "empaquetar" mejor en los tejidos de reserva de los animales (que frecuentemente se desplazan).
Por contra, en el caso de los polisacáridos, el agua de hidratación puede constituir hasta las dos terceras partes de su peso total (por ello les hace más indicados como reserva en los vegetales).
Para las semillas (que deben ser dispersadas), sin embargo, es más importante el peso, por lo que muchas de ellas guardan su reserva energética en forma de aceites (girasol, cacahuete, soja, ricino….)
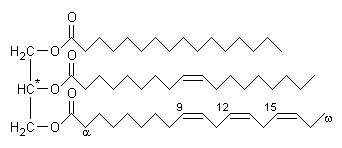
Como recordaréis (ver lípidos), los acilgliceridos constan de una molécula de glicerina esterificada por uno, dos o tres ácidos grasos.
Triglicérido
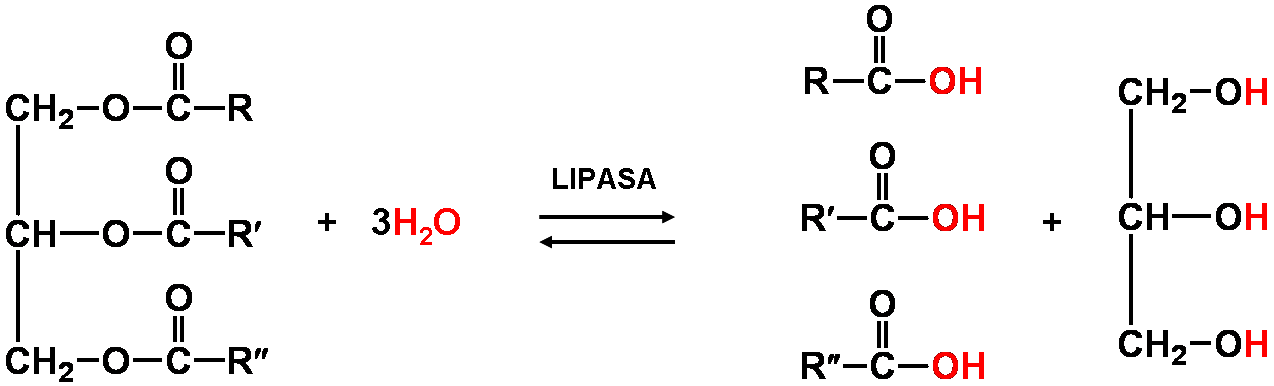
Para poder entrar en la célula, los triacilgliceroles (y los fosfolípidos) deben ser hidrolizados por enzimas denominadas lipasas (lipasa gástrica, lipasa pancreática) que rompen el enlace éster, obteniéndose glicerina y ácidos grasos.
TRIGLICERIDO
GLICERINA + 3 ACIDOS GRASOS
FOSFOLIPIDO
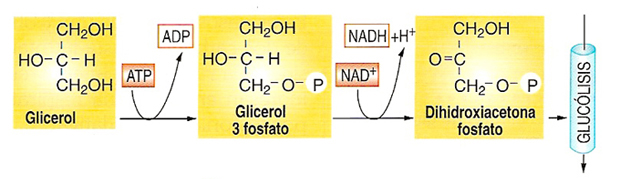
La glicerina (glicerol) que se obtiene de la hidrólisis de muchos lípidos puede transformarse en DHA 3P (dihidroxiacetona 3 P), integrándose en la segunda fase de la glucolisis y degradarse totalmente en el ciclo de krebs.
1) Transformación de la glicerina (o glicerol) en glicerol 3 P a través de la glicerolquinasa con el gasto de 1 ATP.
2) Oxidación del glicerol 3 P y formación de la Dihidroxiacetona 3 P mediante la glicerol fosfato deshidrogenasa con la colaboración de NAD y formación de NADH + H+. La dihidroxiacetona fosfato se puede incorporar a la glucolisis en el 5º paso y seguir la respiración celular hasta el final, o producir glucosa por gluconeogénesis.
La respiración aerobia de una molécula de glicerol puede proporcionar hasta 22 ATP.
Los ácidos grasos deben atravesar la membrana mitocondrial y degradarse en su interior.
Veamos el proceso completo de oxidación de los ácidos grasos:
DEGRADACIÓN DE LOS ÁCIDOS GRASOS
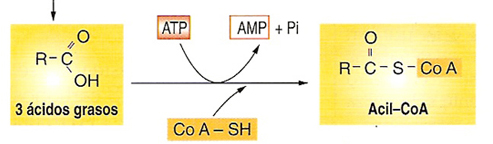
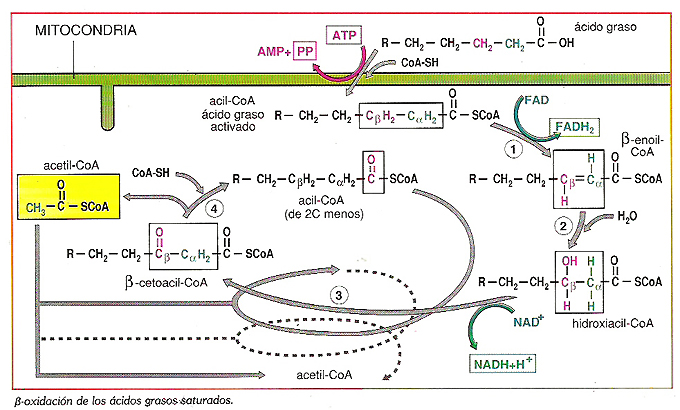
La oxidación de los ácidos grasos tiene lugar principalmente en los peroxisomas de los vegetales y en las mitocondrias de los animales.Para ello deben activarse en forma de Acil CoA, lo cual consume ATP, que se rompe formando AMP + PPi (lo que equivale a 2 ATP).
Ya en la matriz mitocondrial los ácidos grasos serán degradados mediante una ruta llamada ß-oxidación de ácidos grasos porque se va a oxidar el carbono ß del acido graso. En este proceso cada ác. graso se va oxidando, liberándo, paso a paso, "fragmentos" de dos carbonos hasta que termina por consumirse. En cada paso se forman una molécula de FADH2, una de NADH y un acetil-CoA.
Los transportadores de electrones FADH2 y NADH liberan la energía que portan a lo largo de la cadena respiratoria mitocondrial, mientras que el acetil-CoA se incorporará al ciclo de Krebs.
Al final, cada molécula de ácido graso puede aportar un gran número de moléculas de ATP, más o menos dependiendo de su tamaño molecular y de que sea saturado o no (por ejemplo, un ácido esteárico puede producir hasta 146 moléculas de ATP).El proceso comprende tres etapas:
1) Activación del ácido graso por el CoA. El grupo carboxilo del ácido graso es activado por formación de un enlace tioéster con la coenzima A, lo que genera un acil CoA.
En este proceso se requiere la energía que le suministra la hidrólisis de dos enlaces de alta energía del ATP, formando AMP + PPi (lo que equivale a 2 ATP)
2) Transporte: Los ácidos grasos de menos de 12 carbonos, entran libremente en la mitocondria y allí son activados.
Pero los que tienen 14 carbonos o más, tienen que activarse en el lado citosólico de la membrana mitocondrial externa.
El acil CoA ya formado, va a penetrar en la mitocondria mediante un transportador denominado carnitina (transportador de acil-carnitina) hasta la matriz mitocondrial.
3.- Beta-oxidación de los ácidos grasos (Hélice de Lynen)
Es un ciclo recurrente de cuatro pasos
- Primera oxidación por deshidrogenación del acil-CoA, entre los carbonos α y β, proceso catalizado por una deshidrogenasa con FAD, formándose un acil-CoA insaturado y una molécula reducida de FADH2
- Hidratación del acil CoA insaturado, con rotura del doble enlace del enol formado, y formación de un ß-hidroxiacil-CoA de nuevo saturado y con un grupo alcohólico en el carbono ß.
- Segunda oxidación del carbono ß, catalizada por una deshidrogenasa con NAD+ y transformándose en un grupo cetónico dando lugar a un ß-ceto acil-CoA y una molécula de NADH + H+- Rotura del enlace entre los carbonos β y gamma del cetoacil-CoA por interacción del ß-cetoacil CoA con otra molécula de CoA, obteniéndose un acetil-CoA y un acil-CoA que posee 2 carbonos menos que el que inició el ciclo, que reinicia el “ciclo”.
El acetil CoA formado puede seguir la ruta del ciclo de Krebs y degradarse totalmente de forma aeróbica.
El acil-CoA de 2 carbonos menos puede sufrir otra ß-oxidacion y originar un acil-CoA de 2 carbonos menos a su vez (total 4 C menos) y así sucesivamente hasta que solo se obtengan moléculas de acetil-CoA de 2 C, en ese momento habrá completado la llamada hélice de Lynen.
ver animación beta-oxidación de los ácidos grasos
Procesos implicados en la oxidación de un ácido graso
BALANCE ENERGETICO DE LA ß-OXIDACION
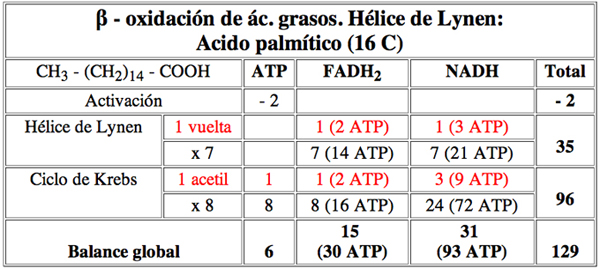
Lo vamos a calcular a partir de un ácido graso, el acido palmítico de 16C
CH3-(CH2)14-COOH
Para que se oxide totalmente ha de dar 7 vueltas de la beta-oxidacion produciendo en total 8 moléculas de acetil-CoA
nº vueltas = (n-2)/2n = número de carbonos del ácido graso
1 A. palmitico + 8 CoASH + 1 ATP + 7 FAD + 7 NAD + 7 H2O
8 acetil CoA + 7 FADH2 + 7 (NADH + H+) + AMP + PPi
Recordando que cada una de las moléculas de acetil-CoA puede ingresar en el ciclo de Krebs con el rendimiento que ello conlleva y que los FADH2 y los NADH pueden penetrar en la cadena respiratoria tenemos que:
8 acetil CoA x 12 ATP (Ciclo de Krebs).................................... 96 ATP
7 FADH2 x 2 ...................................................................... 14 ATP
7 NADH x 3 ....................................................................... 21 ATPTOTAL............... 131 ATP
Si restamos de esa cantidad 2 ATP que se necesita para activar un ácido graso, tenemos que de cada molécula de acido palmítico se obtienen 129 ATP (recordar que la activación del ácido graso requiere la liberación de dos grupos fosfatos de un ATP).
Por tanto: 129 ATP x 7,3 Kcal = 941 Kcal.
Balance de la oxidación del palmítico en términos energéticos
Si el triacilglicérido de partida fuera la tripalmitina obtendríamos 22 ATP de la combustión total de la glicerina y 129 x 3 = 387 de las tres moléculas de ácido palmítico.
En total (3 palmíticos + glicerina) = 387 + 22 = 409 ATP
Rendimiento energético
Energía contenida en un mol de palmítico: 9.790 kJ
Energía almacenada en 129 moles ATP: 129 x 52 kJ = 6.708 kJ
Energía perdida en forma de calor: 9.790 - 6.708 = 3.082 kJ
Eficacia de la síntesis de la ATP a partir del ácido palmítico: 6.708/9.790 = 69%
CATABOLISMO DE PROTEINAS
Como ya vimos cuando las estudiamos, las proteínas poseen otras funciones diferentes que las de reserva energética. Sin embargo sus monómeros o unidades, los aa', pueden ser utilizadas para liberar la energía que contienen los enlaces covalentes entre sus carbonos.
Por ejemplo, un animal carnívoro puede obtener hasta un 90% de la energía que necesita a partir de la oxidación de aminoácidos.
Las plantas en cambio, nunca usan aminoácidos como fuente de energía.
En el caso del ser humanoSe pueden utilizar las proteínas como fuente de energía en casos de ayuno prolongado (o en enfermedades como la diabetes mellitus), aunque esto puede poner en peligro la salud del individuo y si es joven, puede alterar su crecimiento.
También cuando hay un exceso de aminoácidos, debido a una rica rica en proteínas, y se sobrepasa las necesidades corporales para la síntesis de proteínas, y ya que no se pueden almacenar ni excretar, son utilizados como fuente de energía.
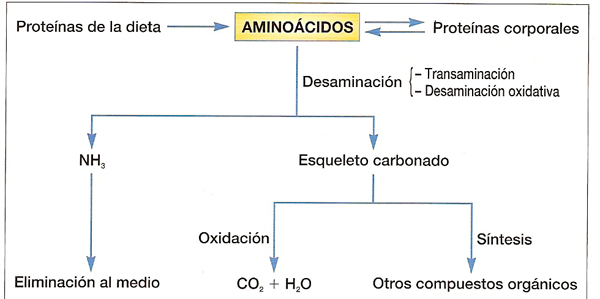
Una vez hidrolizadas las proteínas en el tubo digestivo mediante enzimas proteasas, los aa' resultado de la hidrólisis ingresan en las células y pueden sufrir diferentes tipos de oxidación que determinarán la entrada de sus derivados en el ciclo de Krebs a través de varios puntos, por ejemplo en forma de ácido pirúvico.
Los aminoácidos resultantes de la descomposición de las proteínas se degradan en tres fases:- primero se separan los grupos amino de los aminoácidos
- posteriormente, se oxida el "esqueleto carbonado" que quede.
- finalmente, se eliminan los grupos amino y/o amonio a través de la excreción
Etapas en la oxidación de los aa':
a) Separación de los grupos amino: se realiza mediante dos reacciones sucesivas denominadas transaminación y desaminación oxidativa:
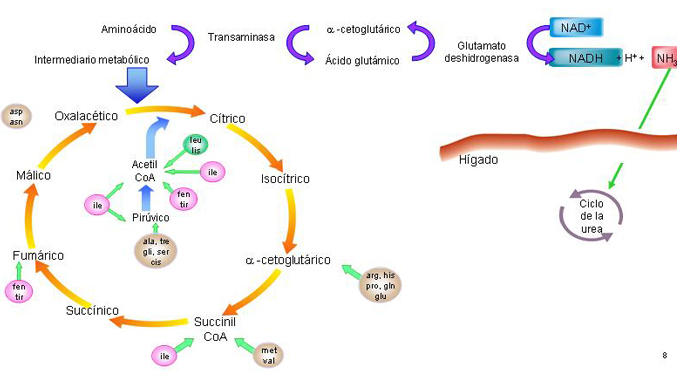
Transaminación
Consiste en el traspaso del grupo amino de un aa' a un alfa-cetoácido (generalmente alfa-cetoglutárico, que se transforma en ácido glutámico). Es decir, un aa' se degrada permitiendo que otro se forme.
Estas reacciones son catalizadas por enzimas transaminasas que se encuentran tanto en el citoplasma como en la matriz mitocondrial de todas las células, especialmente las hepáticas.
Desaminación oxidativa
Es la liberación directa de los grupos amino de los aminoacidos (principalmente del ácido glutámico) y su liberación al medio en forma de NH4+.El glutamato formado por las transaminasas llega a las mitocondrias del hígado, donde el grupo amino se libera en forma de ión amonio. Así se regenera el ácido alfa-cetoglutárico.
Esta reacción es catalizada por deshidrogenasas específicas (en el caso del glutámico, la glutamato deshidrogenasa) con intervención del NAD u otras coenzimas. Tiene como inhibidores al GTP y al ATP y como activadores el GDP y el ADP. De esta forma, la situación energética de la célula inhibe o activa esta vía.
Estas deshidrogenasas se encuentran en el citoplasma y en las mitocondrias hepáticas.
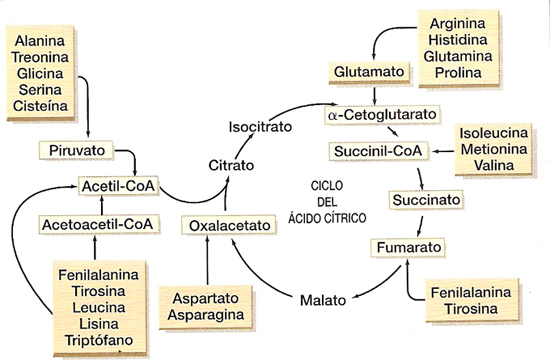
b) Destino de los esqueletos carbonados (transformación del resto resultante en ácido pirúvico, acetil-coenzima A o en algún compuesto del ciclo de Krebs).
Si el organismo no necesita energía, a partir de los aminoácidos puede sintetizarse glucosa o ácidos grasos, pero también pueden seguir la ruta catabólica y transformarse en Acetil-CoA o en ciertos intermediarios del ciclo de Krebs. En cualquier caso, a través del ciclo de Krebs se oxidan totalmente a CO2 y H2O.
Según el tipo de aminoácido se transformará en un compuesto u otro y entrará al ciclo de Krebs (depende de cada aminoácido existen diferentes puntos de entrada).
De esta manera, la cadena carbonada será completamente oxidada.
Vías de entrada de los diferentes aminoácidos al ciclo de Krebs
En ocasiones, el acetil coA formado en el hígado durante la degradación de los aminoácidos en ácidos grasos, se convierte en cuerpos cetónicos, como la acetona, que se exportan a otros tejidos como combustible
c) Eliminación de los grupos aminos y excreción del nitrógeno amónico
Los grupos aminos liberados forman amoniaco que en el agua dan lugar al ión amonio.
El ión amonio es muy tóxico ya que aumenta el ph, por lo que normalmente ha de ser eliminado o transformado en otras sustancias como la urea. El amoníaco es una sustancia tóxica que muchos animales, como los mamíferos, lo empaquetan en el hígado en forma de urea mediante un proceso conocido como ciclo de la urea. La urea pasa al torrente sanguíneo, llega a los riñones y se excreta a través de la orina.
Reacción de formación de la urea
Los animales amoniotélicos excretan el amoniaco directamente al agua a través de las branquias fundamentalmente.
En este grupo se incluyen muchos invertebrados acuáticos y la mayoría de los peces.
En los ureotélicos, el amoniaco es transformado en urea mediante un proceso cíclico denominado ciclo de la urea que tiene lugar en el hígado.La urea es muy soluble y se transporta por la sangre hasta el riñón.
Son ureotélicos algunos peces, como el tiburón, los anfibios terrestres (los renacuajos excretan amoniaco) y los mamíferos.
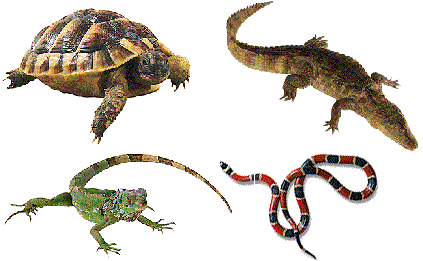
Los uricotélicos convierten el amoniaco en ácido úrico, insoluble en agua, por lo que se excreta en forma de una pasta semisólida.
Los representantes más importantes de estos animales son las aves, los insectos y los reptiles.
Esquema del catabolismo de aminoácidos (proteínas)
Visión general del catabolismo de los aminoácidos
CATABOLISMO DE ACIDOS NUCLEICOS
Los acidos nucleicos se degradan en el tubo digestivo de los animales en sus unidades nucleotídicas (por enzimas nucleasas de la mucosa duodenal y pancreática de los vertebrados).
Posteriormente, otras enzimas, descomponen los nucleótidos en sus grupos fosfatos, pentosas (ribosa o desoxirribosa) y en bases nitrogenadas gracias a la presencia de enzimas nucleasas en la mucosa duodenal de los vertebrados.
Las pentosas siguen la vía de los glúcidos en su degradación
El acido fosfórico se excreta en parte como ión fosfato en la orina y en parte se utiliza para la sintesis de ATP y de nuevos acidos nucleicos (replicación)
Las bases nitrogenadas o bien se utilizan para la síntesis de nuevos nucleótidos o bien experimentan un proceso degradativo hasta acido úrico, urea o amoniaco que serán posteriormente excretados. Este proceso es diferente según sean bases puricas o pirimidinicas.- El catabolismo de las bases púricas conduce a la formación de acido urico.
En algunos animales este acido es excretado directamente, mientras que en otros se transforma en alantoina, en acido alantoico, urea o amoniaco. Las arañas pueden eliminar guanina sin catabolizar.
- Las bases pirimidínicas son catabolizadas mediante un proceso algo diferente pero en definitiva se pueden transformar en urea y amoniaco.
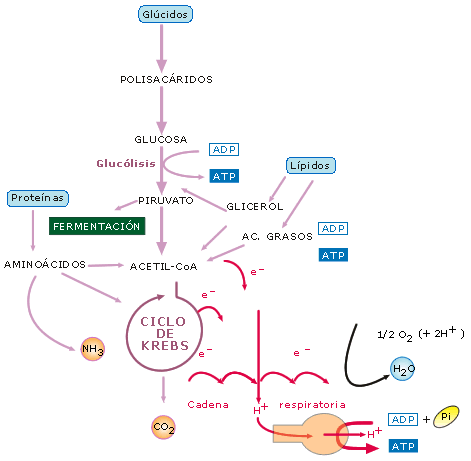
ESQUEMA GENERAL DEL CATABOLISMO
ejercicio on-line sobre catabolismo