ANABOLISMO III
FOTOSÍNTESIS
(continuación)
MODELOS FOTOSINTÉTICOS DE PLANTAS: PLANTAS C3, C4 Y CAM
En el reino Vegetal pueden distinguirse tres grandes grupos de plantas con características fisiológicas, morfológicas y bioquímicas específicas, con un proceso fotosintético diferente en cada grupo.
Plantas C3Las plantas C3, son las que hemos visto, las que fijan el dióxido de carbono a través de la ribulosa difosfato para formar dos moléculas de ácido 3-fosfoglicérico (un compuesto de 3C) mediante el ciclo de Calvin; reacción catalizada por la enzima ribulosa difosfato carboxilasa.
Ejemplos: avena, cebada, arroz, centeno, trigo...
Trigo
Plantas C4 (ciclo de Hatch-Slack)Sin embargo, ciertas plantas de origen tropical como la caña de azúcar o el maíz se denominan plantas C4 porque el CO2 se fija inicialmente al fosfoenolpiruvato dando un compuesto de 4C: el oxalacetato.
Caña de azúcar
Maiz
La enzima que cataliza este proceso se denomina fosfoenolpiruvato carboxilasa (PEPC).
Esta enzima fija el CO2 al fosfoenol piruvato (PEP) dando lugar a oxalacetato.
La fosfoenolpiruvato carboxilasa (PEPC) se halla solo en el citosol de un tipo de células de las hojas denominadas células del mesófilo.
Posteriormente, el oxalacetato formado se convierte en malato o en aspartato, que pasan a las células de la vaina, allí cede el CO2, que es fijado por la RuBisCO.
Las enzimas del ciclo de Calvin se encuentran solo en los cloroplastos de las células de la vaina del haz vascular.
Todo el proceso se llama ruta C4 o ciclo de Hatch-Slack.
Ruta C4 o ciclo de Hatch-Slack
En las plantas C4 la anatomía foliar generalmente
se modifica de manera que se forman dos tipos celulares
implicados en la fotosíntesis: las células de la vaina y las
células del mesófilo.Las células de la vaina con una pared más gruesa, se
distribuyen formando un anillo alrededor de los haces
vasculares y las del mesófilo se encuentran bajo la
epidermis rodeando a las de la vaina.
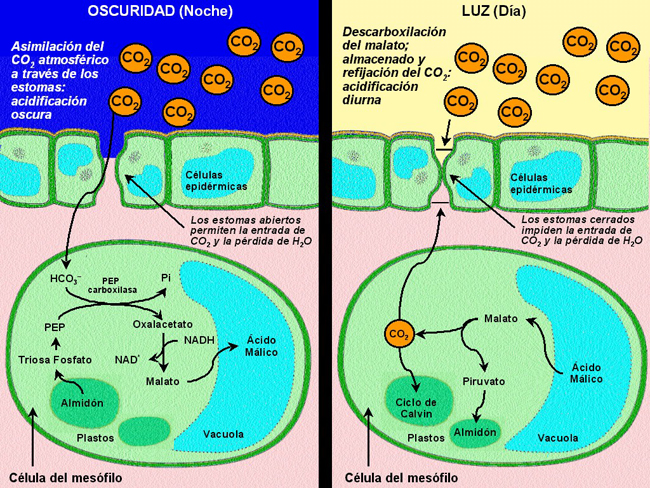
Plantas CAM (Crassulacean acid metabolism)Las plantas CAM fijan el CO2 mediante un proceso modificado del tipo C4, llamado metabolismo ácido de las crasuláceas (la familia de plantas en las que se descubrió por primera vez este proceso. No se halla restringido a ellas).
Plantas CAM
Especies de cactus del género Euphorbia, la piña tropical o la vainilla pertenecen al grupo de las plantas CAM. Algunas de estas plantas tienen grandes células en hojas suculentas, cuyos estomas se abren en la noche para permitir que las enzimas carboxilasas fijen CO2 en compuestos orgánicos con cuatro carbonos.
El metabolismo CAM de plantas originarias de ambientes muy cálidos y secos permite reducir las pérdidas de vapor de agua a través de los estomas.
Es un mecanismo muy similar a la ruta C4, con la diferencia de que la fijación de CO2 por la PEPC y por la RubisCO no están separadas en el espacio, sino en el tiempo.
- Por la noche, cuando el aire es más fresco y húmedo, se abren los estomas y entra el CO2. La PEPC fija el CO2 y forma oxalacetato, que se reduce a malato
- Por el día los estomas se cierran para evitar la pérdida de agua. El malato se descarboxila en el cloroplasto y el CO2 liberado es fijado por la RuBisCO








ver animación plantas C3 y CAM
ver animación comparación plantas C3, C4 y CAM
Diferencias entre las plantas C-3 y C-4
- Las plantas C4 usan más eficientemente el CO2 y además tienen la ventaja adicional de ser más eficientes en el uso del agua que las plantas C3.
- Las plantas CAM son aún más eficientes en el uso del agua que las plantas C4, ya que abren sus estomas solamente de noche, con lo que limitan enormemente la pérdida de agua por transpiración. Estas plantas requieren relativamente menos agua.
- Otra importante diferencia entre las plantas C3 y C4 es que, a una concentración normal de CO2 (0,03%), las C3 alcanzan la saturación fácilmente a bajas intensidades de luz (11.000 a 43.000 lux). Por el contrario, la saturación lumínica es difícil de alcanzar por las C4.
FOTORRESPIRACIÓN (RUTA DEL GLICOLATO o RUTA C2)
Es un proceso que se desencadena como consecuencia de la actividad oxigenasa que tiene la RuBisCo y cuando no hay suficiente CO2 disponible para la planta.
El CO2 no siempre se encuentra disponible para las células fotosintéticas de la planta. Como ya hemos visto, entra en la hoja por los estomas, orificios especializados que se abren y se cierran, dependiendo, entre otros factores de la cantidad de agua. Cuando la planta está sometida a unas condiciones calurosas y secas, debe cerrar sus estomas para evitar la pérdida de agua. Esto provoca también una disminución del CO2 y permite que el oxígeno producido en la fotosíntesis de acumule.
Características de la RuBisCo:a) está considerada como la proteína más abundante de la Tierra, ya que representa el 50% de las proteínas que se encuentran en las hojas verdes.
b) la eficacia fotosintética depende en gran medida de su actividad.
c) es una enzima bifuncional, es decir, según las [CO2] y [O2] puede desarrollar una u otra actividad. Para determinadas concentraciones CO2 / O2, el oxígeno compite con el dióxido de carbono -> Rubisco como oxigenasa > RibulosaBP --> CO2 + H2O.
- Actividad carboxilasa: cataliza la combinación de la ribulosa-1-5-di-P con el CO2.
- Actividad oxigenasa: cataliza la combinación de la ribulosa-1-5-di-P con el O2. Esta oxigenación de la ribulosa-1-5-di-P origina dos moléculas:
- una C3, fosfoglicérico (como en la carboxilación).
- otra C2, fosfoglicocólico o fosfoglicolato (las transformaciones de este ácido constituyen la vía metabólica de la fotorrespiración).
Comparación entre el proceso fotorrespiratorio y el proceso fotosintético
Esta reacción es el primer paso de un proceso conocido como fotorrespiración, por el cual los glúcidos son oxidados a CO2 y agua, en presencia de luz.
El fosfoglicolato se desfosforila en el cloroplasto a glicolato, producto sin utilidad metabólica que en algunas algas se excreta al medio. Pero normalmente, el glicolato se recicla mediante un proceso conocido como la ruta del glicolato o ruta C2, en el que además de los cloroplastos, participan también los peroxisomas y las mitocondrias.
El proceso convierte dos moléculas de glicolato (4 carbonos) en una de 3 fosfoglicerato (3 carbonos) y una de CO2.
Orgánulos implicados en la fotorrespiración
ver animación fotorrespiración
Características de la fotorrespiración
a) es una vía metabólica que se produce al mismo tiempo que la fotosíntesis, cuando el ambiente es cálido y seco (los estomas de las hojas se cierran para evitar la pérdida de agua por transpiración), por tanto, se produce en presencia de luz.
b) implica un consumo de O2 porque la RuBisCo actúa como oxigenasa (y oxida a la ribulosa-1-5-di-P para transformarla en ác. 3-P-glicérico y ác. P-glicocólico).
c) el ác. P-glicocólico pasa a los peroxisomas, donde por cada dos moléculas del mismo se obtiene una de P-glicérico y una de CO2.
d) La combinación de la actividad oxigenasa de la RuBisCO y la ruta del glicolato, que tiene lugar en condiciones de ilumnación de las plantas, consume O2 y genera CO2. Por este motivo se denomina fotorrespiración, aunque a diferencia de la respiración mitocondrial, no conserva energía en forma de ATP sino que la consume.
e) es un mecanismo que disminuye la eficacia fotosintética de un 30 a un 50% y además consume ATP. Esta pérdida de carbono fijado, en forma de CO2 liberado por fotorrespiración, representa un lastre para la planta, ya que consume materia orgánica ya formada sin producir ATP, es decir, deshace parte de lo conseguido en la fotosíntesis.
A diferencia de la respiración mitocondrial, la fotorrespiración es un proceso donde la energía se pierde, y no se produce ni ATP ni NADH.
El papel de la fotorrespiración sigue sin conocerse con certeza ya que limita la eficacia de la fotosíntesis, quizás protección frente a fotooxidación. Es posible que se trate de una vía relíctica heredada de tiempos geológicos en los que la relación CO2/O2 de la atmósfera era mayor que la actual.
ver reacciones fotorrespiración
FACTORES QUE INFLUYEN EN EL PROCESO FOTOSINTÉTICO
El rendimiento de la fotosíntesis puede ser medido fácilmente por la cantidad de CO2 absorbido por la planta. En él influyen:
La intensidad y longitud de onda de la luzCada especie se encuentra adaptada a desarrollar su vida dentro de un intervalo de intensidad de luz, por lo que existirán especies de penumbra (esciófilas) y especies fotófilas
Dentro de cada intervalo, a mayor intensidad luminosa, mayor rendimiento, hasta sobrepasar ciertos límites, en los que sobreviene la fotooxidación irreversible de los pigmentos fotosintéticos.
Para una igual intensidad luminosa, las plantas C4 (adaptadas a climas secos y cálidos) manifiestan un mayor rendimiento que las plantas C3, y nunca alcanzan la saturación lumínica.
El color de la luzYa sabemos que los carotenos y las clorofilas de los fotosistemas absorben fotones de una determinada longitud de onda. Por lo tanto, si se ilumina una planta con luz de longitud de onda inadecuada o con una intensidad insuficiente, la fotosíntesis no podrá realizarse y la planta no se desarrollará.
La clorofila α y la clorofila β absorben la energía lumínica en la región azul y roja del espectro, los carotenos y xantofilas en la azul, las ficocianinas en la naranja y las ficoeritrinas en la verde. Estos pigmentos traspasan la energía a las moléculas diana. La luz monocromática es menos aprovechable en los organismos que no tienen ficoeritrinas y ficocianinas.
En las cianofíceas, que también poseen estos pigmentos anteriormente citados, la luz roja estimula la síntesis de ficocianina, mientras que la verde favorece la síntesis de ficoeritrina.
El tiempo de iluminaciónExisten especies que desenvuelven una mayor producción fotosintética cuanto mayor sea el número de horas de luz, mientras que también hay otras que necesitan alternar horas de iluminación con horas de oscuridad.
La escasez de aguaAnte la falta de agua en el terreno y de vapor de agua en el aire, disminuye el rendimiento fotosintético. Esto se debe a que la planta reacciona ante la escasez de agua cerrando los estomas para evitar su desecación, dificultando de este modo la penetración de dióxido de carbono.
Además, el incremento de la concentración de oxígeno interno desencadena la fotorrespiración.
Este fenómeno explica que en condiciones de ausencia de agua, las plantas C4 sean más eficaces que las C3.
TemperaturaLa fotosíntesis, como todo proceso químico, está influenciada por la temperatura, ya que por cada 10° C de aumento de temperatura, la velocidad del rendimiento fotosintético se duplica.
Ahora bien, un aumento excesivo de la temperatura desnaturalizará las enzimas que catalizan el proceso y se producirá un descenso del rendimiento fotosintético.
Concentración de CO2Si el resto de los factores se mantiene constante, un aumento en la cantidad de CO2 existente aumentará el rendimiento de la fotosíntesis hasta llegar a un valor máximo por encima del cual se estabilizará.
Concentración de O2
Un aumento en la concentración de O2 inhibe la fotosíntesis, ya que el oxígeno inhibe la enzima que incorpora el CO2 a la Ribulosa-1-5-difosfato (RuBP).
FIJACIÓN FOTOSINTÉTICA DEL N Y S
(REDUCCIÓN DE NITRATOS Y SULFATOS)En la fotosíntesis, además de glucosa, se elaboran otros compuestos (aminoácidos y nucleótidos), que contienen grupos amino (-NH) y tiol (-SH).
Tanto en el agua y en el suelo, el N y el S se encuentran en forma de compuestos oxidados (NO3- y SO4=), que han de ser absorbidos y reducidos por las plantas para ser incorporados a la materia orgánica.
En ambos casos, el proceso es lineal (no cíclico) y se sirve del NADPH2 generado en la fase luminosa de la fotosíntesis.
La reducción fotosintética del NLas plantas pueden obtener el nitrógeno que necesitan a partir de los nitratos (NO3-) por ejemplo. Los nitratos son absorbidos por las raíces y transportados por los vasos leñosos hacia el parénquima clorofílico de la hoja.
En los nitratos, el nitrógeno se encuentra en una forma muy oxidada, mientras que en los compuestos orgánicos se encuentra en forma reducida. La reducción es realizada por el NADPH2 y la energía necesaria para el proceso es aportada por el ATP.
Ambos productos, como ya sabemos, se obtienen en grandes cantidades en la fase luminosa de la fotosíntesis. Esta es la razón por la que la reducción del nitrógeno y su incorporación en las sustancias orgánicas se realiza en los cloroplastos, y no porque el proceso necesite de una manera directa la luz.
Comprende dos etapas catalizadas por enzimas específicas:
1) Etapa 1: La transformación de nitratos en nitritos y de éstos en amonio (recordar ciclo del N en CTMA).
2) Etapa 2: El amonio se combina con el ácido alfa-cetoglutárico para formar glutámico (también el NH3 se combina con el ác. glutámico y forma glutamina, la cual puede transferir grupos amino -mediante transaminasas- a otros compuestos para sintetizar aminoácidos y nucleótidos).
Es el glutámico el aminoácido que servirá posteriormente para donar el nitrógeno a aquellas moléculas orgánicas que lo precisen.
La reducción fotosintética del S
El azufre es absorbido por las raíces en forma de sulfatos (SO4-2) u otras sales y, una vez reducido, es incorporado en otras sustancias orgánicas de una manera similar a la que hemos visto con el nitrógeno (recordar ciclo del azufre en CTMA)
También es un proceso lineal en el que los sulfatos se reducen a sulfitos y éstos a H2S. También requiere NADPH2 y ATP. Una vez reducido, el H2S se puede incorporar como grupo tiol (-HS) a la cisteína
La reducción fotosintética del P
El fósforo es absorbido a partir del suelo por las raíces en forma de fósfato (no necesita reducirse ya que se encuentra en el mismo estado de oxidación en las moléculas orgánicas) y se incorpora al ATP por reacción con el ADP.
FOTOSÍNTESIS EN BACTERIAS
Además de las plantas y las algas, muchas bacterias pueden utilizar la luz para obtener energía. Sin embargo su diversidad metabólica es mucho mayor que la de los eucariotas fotosintéticos.
Se pueden distinguir dos tipos de fotosíntesis:
Fotosíntesis oxigénica
La llevan a cabo las cianobacterias y usan agua como donador de electrones, que al fotooxidarse produce O2, como en las plantas.
Su citoplasma incluye tilacoides, cuyas membranas poseen ambos fotosistemas I y II.
Tienen una sola forma de clorofila (a) y además pigmentos accesorios: ficobilinas.
Un tipo de ficobilina, la ficocianina, es azul y junto con el verde de la clorofila son los responsables del color azul-verdoso de estas bacterias.
Pueden contener además ficoeritrina (de color rojo).
Las cianobacterias reducen el CO2 por el ciclo de Calvin y utilizan nitratos o amonio como fuente de nitrógeno (algunas pueden fijar el N2 atmosférico).
Las cianobacterias fueron los primeros seres fotótrofos oxigénicos y el resultado de su actividad fue la paulatina transformación de la primitiva amósfera anaerobia y reductora a una aerobia y oxidante (la actual).
Cianobacterias
Fotosíntesis anoxigénicaLa realizan las bacterias fotosintéticas.
Las bacterias únicamente son poseedoras de un solo fotosistema y como pigmento fotosintético, bacterioclorofila.
No desprenden oxígeno ya que no utilizan agua como donador de electrones.
Utilizan compuestos de azufre o hidrógeno como donadores de electrones para obtener poder reductor (en forma de NADH), y no para obtener ATP.
Obtienen ATP a través de la fotofosforilación cíclica.
Se realiza exclusivamente en condiciones anóxicas: no solo no generan oxígeno, sino que no lo toleran (son anaerobias estrictas).
Tienen una fase oscura análoga a la de las plantas y cianobacterias
En función de la molécula que emplean como dador de electrones y el lugar en el que acumulan sus productos, es posible diferenciar dos tipos de bacterias fotosintéticas:
Bacterias rojas o púrpuras del azufreHabitan en zonas anóxicas de lagos bien iluminados.
Poseen un solo tipo de fotosistema que se localiza en los pliegues de la membrana plasmática que contiene bacterioclorofila a y b, así como distintos carotenoides responsables de su color púrpura o marrón.
Se caracterizan por emplear sulfuro de hidrógeno (H2S) como donador de electrones.
Acumulan el azufre resultante en su interior.
Bacterias púrpuras del azufre
Bacterias verdes del azufre
Se hallan en las zonas ricas en azufre de los lagos.
También son anaerobias
Solo poseen un fotosistema y los pigmentos fotosintéticos se reúnen en los clorosomas, vesículas membranosas libres en el citoplasma que contienen las bacterioclorofilas.
Su dador de electrones es un compuesto sulfuroso (H2S o S) o hidrógeno molecular (H2) para el paso final de la fase luminosa en la que se reduce NAD a NADH.
En contraste con las purpúreas, no acumulan azufre en su interior, su metabolismo forma deposiciones de sulfato en el exterior de las células
Además poseen vesículas de gas para mantenerse a una profundidad óptima de luz y H2S, puesto que estas bacterias no son móviles.
Utilizan una cadena inversa de electrones inversa para fijar el CO2.
Muchas de estas bacterias participan en el ciclo del azufre
Colonias de bacterias verdes, creciendo en un lago sulfuroso
vídeo fotosíntesis bacteriana con detalle
QUIMIOSÍNTESIS COMO OTRA FORMA DE NUTRICIÓN AUTÓTROFA
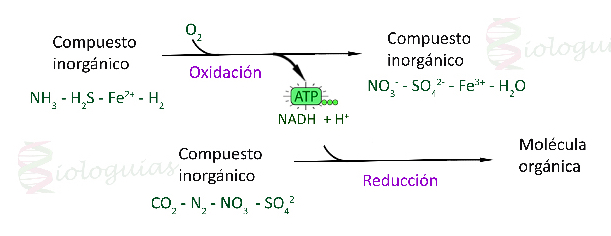
La quimiosíntesis es también una forma de nutrición autótrofa en la que, a diferencia de la fotosíntesis, la energía y los electrones (ATP y NADPH) necesarios para los procesos de anabolismo van a proceder de la oxidación de sustancias inorgánicas.
Se trata de una forma de nutrición típicamente bacteriana: bacterias quimiolitotrofas y el proceso recibe el nombre de quimiosíntesis.
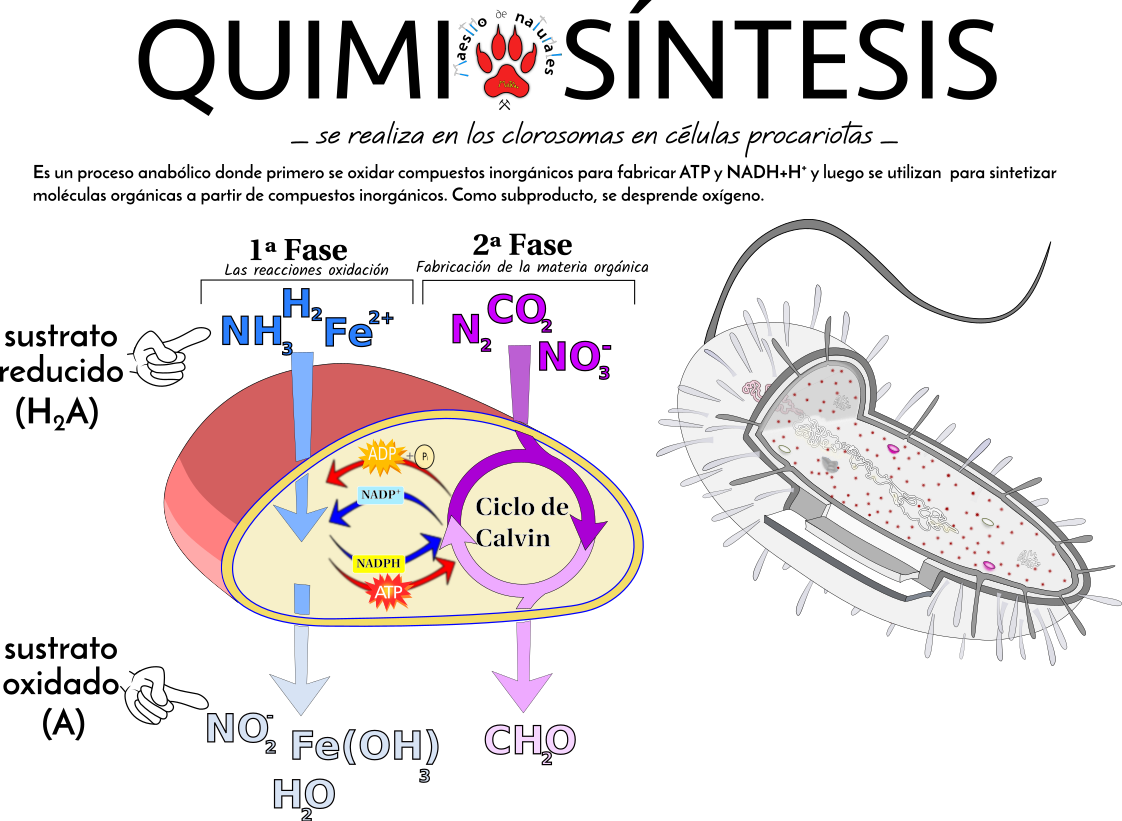
Fases de la quimiosíntesis
Fase oxidativaEn esta fase, un compuesto inorgánico se oxida.
Dependiendo de la especie de bacteria y del sustrato disponible (amoniaco, nitritos, azufre, hiero...) será un compuesto inorgánico u otro.
La oxidación de estos compuestos libera energía en forma de ATP y se genera poder reductor en forma de NADH, de modo análogo a lo visto en la fase luminosa de la fotosíntesis.
Fase de biosíntesis
Esta fase sería análoga a la fase oscura de la fotosíntesis y en ella se fija CO2 (se reduce) y se forma materia orgánica gracias al ATP y NADH obtenidos en la fase anterior
Tipos de quimiosíntesis
Según el sustrato oxidado tendremos:
a) Bacterias nitrosificantes
Como las del género Nitrosomonas que obtienen energía en forma de ATP y coenzimas reducidas por medio de la oxidación del amoniaco (NH4+) a nitrito
Presentes en los excrementos y en la materia orgánica en descomposición.
b) Bacterias nitrificantesComo las del género Nitrobacter que oxidan los nitritos (NO2-) a nitratos (NO3-), que puede ser absorbido nuevamente por las plantas, cerrándose así el ciclo del nitrógeno
c) Bacterias incoloras del azufre
Estas bacterias oxidan generalmente el ácido sulfhídrico (H2S) en presencia de O2, los sulfuros a azufre y el azufre a sulfitos o a sulfatos.
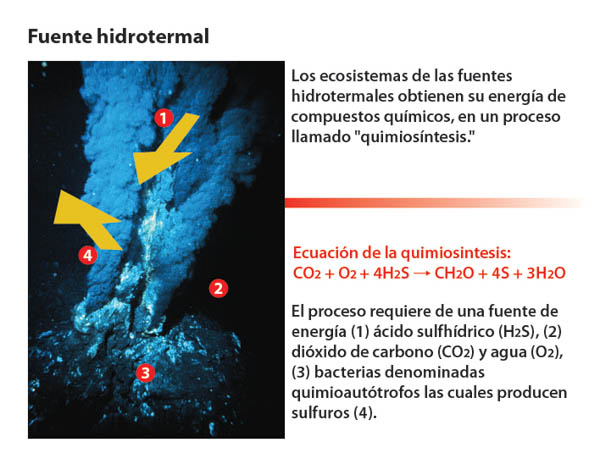
Son organismos típicos de las aguas residuales y emanaciones hidrotermales.
d) Bacterias del hierro
Oxidan los compuestos ferrosos a férricos.
Estos dos últimos tipos de bacterias medran, sobre todo, en los yacimientos de azufre y hierro de origen volcánico y en particular en los llamados humeros negros.
Es de destacar, que las bacterias quimiosintéticas son los únicos seres vivos no dependientes, ni directa ni indirectamente, de la luz solar.
Chimeneas hidrotermales o humeros negros (y blancos)
en las dorsales oceánicas
e) Bacterias del hidrógeno
Utilizan cono dador de electrones el H2. El H2 cede electrones a la cadena respiratoria hasta el O2, liberando agua y energía o bien para reducir directamente el NAD.
También a partir de CH4 -> H2O
Quimiosíntesis: Bacterias quimiolitotrofas.
Energía de oxidación de mol. inorgánicas -> ATP, NADPH (o NADH).
Importantes en los ciclos de la materia en el ecosistema.Bacterias del nitrógeno: Energía de la oxidación: NH4+ -> NO2 -> NO3 -
B. del azufre: S0 , H2S -> SO4 2-
B. del hierro: Fe 2- -> Fe 3-
B. del hidrógeno: H2 , CH4 -> H2O